Die Mantelfärbung deutscher Heringsmöwen Larus fuscus
Andreas Noeske
Andreas Noeske

Heringsmöwe (© Andreas Noeske)
ABSTRACT Between 17th May and 15th June 1986 62 adult Lesser Black-backed Gulls Larus fuscus were caught in a breeding colony on Amrum (54° 42' 21.61" N, 08° 21' 4.00" E) situated in the German Wadden Sea of Schleswig-Holstein. Mantle colour was assessed visually under laboratory conditions by comparing feathers taken from the scapulars and greater coverts with the Munsell Neutral Value Scale and Kodak Grayscale.
The variation according to Munsell ranges from 1.75 to 3.5 (arithmetic mean = 2.85, median = 3.0, modal value = 3.0; average of all three values = 2.95). The variation according to Kodak ranges from 9.7 to 16 (arithmetic mean = 12.1, median = 11.3, modal value = 11.0, average of all three values = 11.5). 46 birds (= 74.2%) show grey values between 11 and 13 typical of intermedius. 18 birds (= 29.0%) show a grey value of 13 or darker which is within the range of fuscus. Eleven gulls (= 17.7%) show a grey tone of 14 or darker which is close to the mean value of fuscus.
Variation, distribution and mean value of mantle coloration characterize the Amrum gulls as intermedius-birds. This is in accordance with the mitochondrial-genetic population structure found by Liebers & Helbig (2002) as the haplotype composition of Amrum Lesser Black-backed Gulls is close to intermedius from southern Norway. In standard body measurements the birds from the Schleswig-Holstein coast resemble graellsii and intermedius closest (Noeske 1989). One bird (Arnhem 5105925, Munsell 3.0, Kodak 11.7) was ringed as nestling on Terschelling, Holland on 12th July 1976. This bird as well as the high proportion of dark-mantled gulls indicate the influence of Dutch and southern Scandinavian birds on the colonies of the German North Sea coast.
Clearly distinctive was a black-mantled gull with a Munsell grey value of 1.75 (Kodak 16). This individual belongs to the darkest Lesser Black-backed Gulls ever recorded. Such black-mantled, long-winged and small Lesser Black-backed Gulls have been widely considered as North-Scandinavian or Baltic fuscus-birds. Such adult birds found in intermedius-colonies might be immigrants from the fuscus range but the bulk of such dark-mantled birds is more likely to have hatched within the intermedius-range and such individuals might be indistinguishable from fuscus all year round. The gene flow across subspecies "boundaries" defined by taxonomists results in a mixture of characters so that neither one single character nor a multiple character approach is diagnostic of fuscus. The criteria for identifiying adult fuscus in late August/September given by Jonsson (1998) are regarded as insufficient in a western European context.
Due to problems with the separation of fuscus from intermedius the Dutch Rarities Committee CDNA has removed all former claims of 'Baltic Gull' after review. Now only ringed fuscus of proven provenance are considered acceptable, i.e. birds ringed as chick on the breeding grounds (van der Vliet et al.). The British Birds Rarities Committee BBRC is in full agreement with the CDNA that on current knowledge fuscus cannot be identified in the field as a vagrant in western Europe although Jonsson's criteria may well be applied in the Baltic region (Colin Bradshaw pers. comm.).
The range of mantle colour given for intermedius by Jonsson (1998), Malling Olsen & Larsson (2003) and Howell & Dunn (2007) does not describe the variation in intermedius-colonies properly. Also the overlap in mantle colour between fuscus and intermedius given by Jonsson (1998) is not representative. The variability of the Amrum Lesser Black-backed Gulls is in full accordance with the findings by Barth (1966, 1968) and the description of intermedius given by Glutz von Blotzheim & Bauer (1982).
Spekulationen über die Zuordnung der im Bereich der Deutschen Bucht brütenden Heringsmöwen Larus fuscus reißen bis heute nicht ab, denn die Population der Küsten zwischen Dänemark und Spanien ist in Bezug auf das Hauptunterscheidungsmerkmal Mantelfärbung nicht einheitlich. In den Kolonien der Niederlande und Deutschlands werden seit langem Vögel mit auffällig hellem und sehr dunklem Mantel beobachtet. Dies gab Anlass zu Vermutungen über deren Herkunft aus Brutgebieten sowohl der westlichen/britischen als auch der intermediären südskandinavischen Population und sogar der schwarzmanteligen Nominatform. Dabei wirkt die Anwendung von Unterartnamen auf hell- und dunkelgraue sowie schwarzmantelige Heringsmöwen durch Vogelbeobachter heute noch oftmals übermäßig selbstsicher, hilfesuchend oder sogar ahnunglos. Dies hat selbst bei Autoren in Verbindung mit zum Teil irrigen Vorstellungen vom Aussehen der drei in Mitteleuropa auftretenden Unterarten zu einem verwirrenden Kaleidoskop an Meinungen über das Vorkommen dieser Möwen geführt (Tinbergen 1929, van Dobben 1931, Goethe 1951, 1957, Barth 1966, 1968, 1975, Hausmann 1969, Busch 1974, Hanoldt 1974, Glutz von Blotzheim & Bauer 1982, Meier & Todt 1985, Zang et al. 1991, Liebers 2000, Svensson et al. 2000, Liebers & Helbig 2002, Wernham et al. 2002, Pfeifer 2003, Bauer et al. 2005, AERC ohne Datum). Ursache hierfür ist die jahrelange Unklarheit über die taxonomische Stellung von intermedius sowie die z.T. vereinfachten Darstellungen in Bestimmungsbüchern in Zusammenhang mit falschen Vorstellungen vom Unterartbegriff.
 |
Abb. 1: Heringsmöwe Larus fuscus, Helgoland, 5. Juni 2007 (© Stefan Pfützke).
Vergleiche mit http://cyberbirding.uib.no/photo/l_fuscus_04.php und http://www.pbase.com/arieouwerkerk/image/58882008
 |
Abb. 2: Heringsmöwe Larus fuscus, Helgoland, Juni 2006 (© Gabriel Schuler).
 |
Abb. 3: Heringsmöwe Larus fuscus, Helgoland, Juni 2007 (© Axel Halley).
 |
Abb. 4: Heringsmöwe Larus fuscus, Helgoland, 9. Juni 2006 (© Sven Achtermann).
Vergleiche mit http://www.wildbirdgallery.com/images/birds/larus_fuscus/graellsii.htm
 |
Abb. 5: Heringsmöwe Larus fuscus, Helgoland, Juni 2007 (© Axel Halley).
Siehe auch http://www.netfugl.dk/pictures.php?id=showpicture&picture_id=10757
Busche & Berndt (1982) wiesen bereits frühzeitig darauf hin, dass mit dem wachsenden Brutbestand in Schleswig-Holstein der Unterartfrage dringend nachgegangen werden sollte. So gehen Malling Olsen & Larsson (2003: 372) immer noch davon aus, dass der subspezifische Status im Bereich südwestliches Dänemark, Schottland und Niederlande unsicher ist. Glutz von Blotzheim & Bauer (1982) ließen deshalb die Frage nach der Unterartzugehörigkeit offen, da vor der wahrscheinlich richtigen Zuordnung zu intermedius eine eingehende Untersuchung der westeuropäischen Festlandpopulation von Westjütland bis Spanien angezeigt wäre. Nach Haffer (1985) sind Subspezies-Namen als taxonomisches Hilfsmittel durchaus nützlich und vertretbar, wichtiger aber ist eine statistisch-quantitative regionale Analyse der geographischen Variation mit möglichst genauer kartographischer Darstellung der Ergebnisse. Eine auf Nachvollziehbarkeit aufbauende sachliche Beurteilung der Mantelfärbung zusammen mit neuen Erkenntnissen aus der molekularen Populationsgenetik des Silber-Heringsmöwen-Komplexes Larus argentatus/fuscus bleiben hierbei der Schlüssel zum Verständnis der Beziehungen des schleswig-holsteinischen Brutbestandes der Heringsmöwe zu den Nachbarpopulationen im Norden, Osten und Westen. Dieser Beitrag fasst die Ergebnisse zur Mantelfärbung in einer Heringsmöwen-Kolonie auf Amrum zusammen und schließt damit die Datenlücke in der Inneren Deutschen Bucht. Darüber hinaus wird eine Einschätzung abgegeben, inwieweit Vögel mit Merkmalen von fuscus und graellsii in der Deutschen Bucht tatsächlich diesen Taxa zugeordnet werden können.
Material und Methode
Zwischen dem 17. Mai und 15. Juni 1986 wurden im NSG Amrum-Odde (54° 42' 21.61" N, 08° 21' 4.00" E, Google Maps) 62 adulte Heringsmöwen auf dem Nest gefangen und von mir gewogen und vermessen. Für die Grauwertbestimmung wurden von jedem Vogel mehrere Federn aus dem Bereich der Schultern und Großen Flügeldecken gesammelt und in einem Briefumschlag für die spätere Untersuchung aufbewahrt. Eine verläßliche Beurteilung der Mantelfärbung, wie sie für diese vergleichende Darstellung erforderlich ist, ist im Feld bei nicht gefangenen Vögel unmöglich. Bei starker Bewölkung sahen Amrumer Vögel, die weit entfernt standen, meist einheitlich schwarz aus. Bei sehr starker Sonneneinstrahlung wirkten die Vögel durch starke Reflektionen an der Federstruktur meist zu hell. Alle Grauwertbestimmungen wurden daher von mir zu Hause visuell durch den Vergleich mit der Munsell Neutral Value Scale durchgeführt (Glossy, 37-Step Scale, 1971 Edition der Munsell Color Company, Baltimore, Maryland, als Beilage enthalten in Munsell Color Company 1966). Außerdem wurden die Federn mit dem Farbatlas von Munsell (1929) verglichen und fotografisch dokumentiert (Schwarz-Weiß Film Ilford FP 4, geblitzt).
Im März 2007 wurden die vor Lichteinflüssen geschützt aufbewahrten Federn erneut mit einer Munsell Neutral Value Scale verglichen (Matte Edition, 31-Step Scale, Abb.6). Im Gegensatz zu der 1986 verwendeten Ausgabe ist die Grauskala dieses Farbfächers von 2/ bis 9.5/ in viertel Schritten eingeteilt, wodurch eine präzisere Bestimmung möglich ist. Der Fehler ist nach eigener Einschätzung kleiner als +/- 0,25.
 |
Abb. 6: Munsell Neutral Value Scale, Matte Edition, 31-Step Scale, Farbstandards.de.
Bei der visuellen Grauwertbestimmung wurden mindestens 2-3 Federn übereinander gelegt und auf die Grautafeln angedrückt. Durch das Übereinanderlegen von mindestens zwei Federn wurden Verhältnisse geschaffen, die der natürlichen Federlage beim Vogel entsprechen. Außerdem wurde damit verhindert, dass bei einer Feder der durchscheinende Untergrund die Beurteilung beeinflußt. Dabei ist es notwendig, den Betrachtungswinkel so zu verändern, dass Spiegelungen durch die Federstruktur vermieden werden und sich die distale Hälfte der Außenfahne möglichst mit dem Grauton auflöst. Die Bestimmung wurde bei natürlichem Tageslicht ohne direkte Sonneneinstrahlung vorgenommen. Das Munsell-Farbensystem wurde auch von Barth (1966) in seiner grundlegenden Arbeit für die exakte Grauwertbestimmung an Museumsbälgen verwendet.
Darüber hinaus wurde der Grauwert mit der Kodak Grayscale bestimmt, die auch von Jonsson (1998) und Malling Olsen & Larsson (2003) bei Heringsmöwen verwendet wurde. Diese Grauwertbestimmung wurde von Andreas Buchheim, Martin Gottschling und mir gleichzeitig und unabhängig durchgeführt. Bei 38 Vögeln (= 61%) wurde Übereinstimmung in der Beurteilung erreicht (z.B. "11,11,11"), bei 24 Vögeln (= 39%) gab es eine Abweichung (z.B. "9,10,10"). Für die Auswertung wurde der Mittelwert aus diesen drei Beurteilungen gebildet. Andere Beurteilungen wie etwa "10,11,12" oder "13,15,15" gab es nicht. Der Fehler bei dieser dreifachen Beurteilung kann als < 0,5 angesehen werden.
Für die Einstellung der Grauwerte in den Farbtafeln wurde die Mantelfärbung mit der CGI simulation of Kodak Grayscale verglichen und in Photoshop nach dem L- und B%- Wert sowie der Farbsättigung so eingestellt, dass nach meinem Eindruck die Unterschiede zwischen der darzustellenden Mantelfärbung und der Kodak-Skala am geringsten sind, dabei aber ein natürlicher Feldeindruck entsteht, der sich durch die neutralen Grautöne nicht ergibt. In der Hand haben die Federn einen mehr oder weniger deutlichen Braunton, an der Küste dagegen wirkt der Mantel oft leicht blau(grün)stichig. Die Farbsättigung in den Abbildungen wurde den Feldbedingungen und durch den Vergleich mit Fotos angepasst.
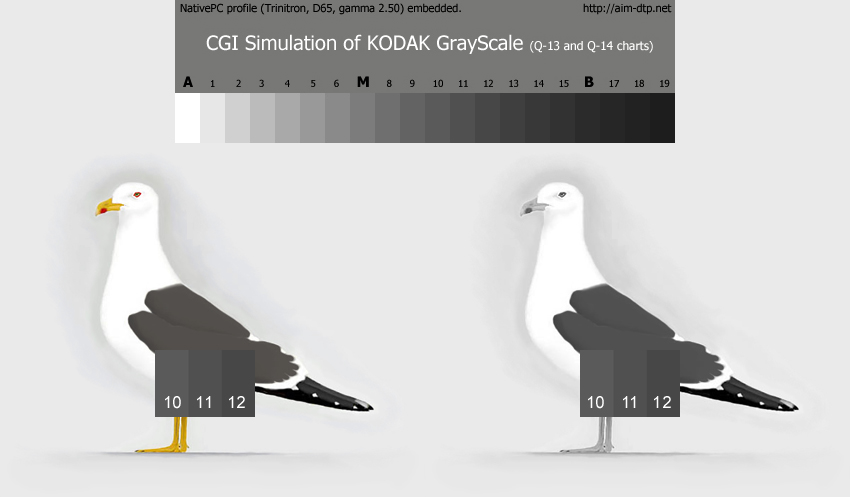 |
Abb. 7: Einstellung der Mantelfärbung in den Farbtafeln für alle Graustufen der Heringsmöwe nach der Kodak Grayscale (© Andreas Noeske).
Ergebnisse
Die geografische Variation ist vor allem in der Mantelfärbung deutlich (Abb.8). Der Mantel ist bei den graellsii-Populationen am hellsten. Die fuscus-Populationen wirken nahezu einheitlich schwarz. Die intermedius-Population Südskandinaviens nimmt eine Mittelstellung ein. Ihre Mantelfärbung zeigt eine besonders hohe Variabilität, so dass in Brutkolonien dieser Unterart sowohl hellgraue als auch schwarze Vögel auftreten, die jeweils den Vögeln der britischen bzw. baltischen Unterart gleichen. Sie ist im Mittel aber sowohl von den britischen Heringsmöwen als auch von den Vögeln der Nominatform deutlich verschieden (Voous 1963, Glutz von Blotzheim & Bauer 1982).
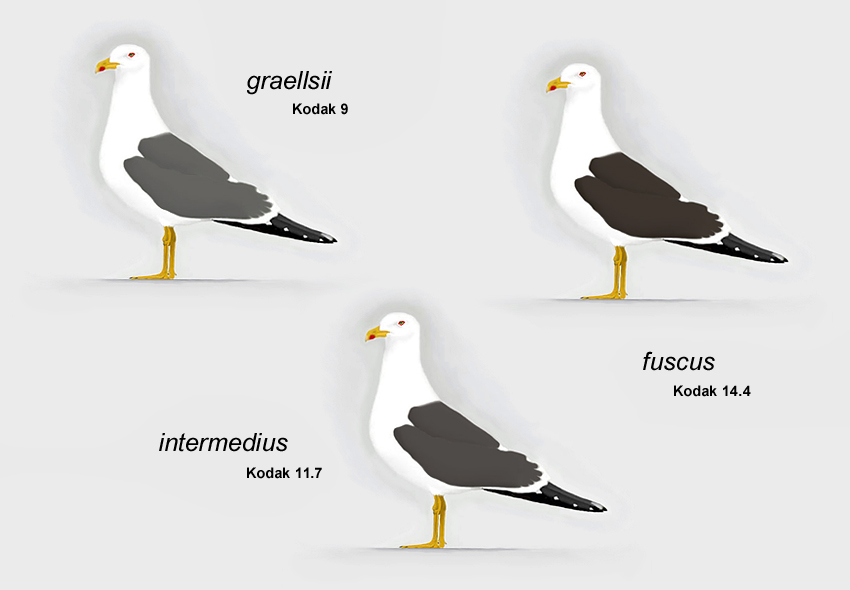 |
Abb. 8: Heringsmöwen Larus fuscus mit Mantelfärbungen, die ungefähr die durchschnittlichen Unterschiede zwischen graellsii, intermedius und fuscus wiedergeben (© Andreas Noeske).
Amrumer Grauwerte nach Kodak
Nach Jonsson (1998) und Malling Olsen & Larsson (2003) haben die drei Unterarten folgende Kodak- Werte:
fuscus 13-17, Mittelwert = 14,4 (13-15 nach Howell & Dunn 2007);
intermedius 11-13, Mittelwert = 11,7;
graellsii 8-10(11) (9-11 nach Howell & Dunn 2007), Mittelwert ~9 für den Süden Englands (Klaus Malling Olsen in litt.).
Amrumer Heringsmöwen variieren von 9,7-16 (Mittelwert = 12,1; Median = 11,3; Modalwert = 11,0, "11,11,11" bei 25 Vögeln = 40%) und weisen damit Grauwerte auf, die sowohl bei graellsii als auch bei fuscus vorkommen. Alle drei Werte ergeben einen "gemittelten Mittelwert" von 11,5. Die große Mehrheit (46 Vögel = 74,2%) weist mit "11,11,11" bis "13,13,13" typische intermedius-Grauwerte auf. 18 Vögel (= 29%) haben einen Kodak-Wert 13 oder dunkler und liegen damit innerhalb der für fuscus beschriebenen Variationsbreite. Elf Vögel (= 17,7%) wurden als "14,14,14" oder dunkler beurteilt und liegen außerhalb der von Jonsson (1998) und Malling Olsen & Larsson (2003) angegebenen Variationsbreite für intermedius. Lediglich ein Vogel (= 1,6%) hatte mit "9,10,10" einen für graellsii typischen Kodak-Wert.

Abb. 9: Variationsbreite bei Amrumer Heringsmöwen Larus fuscus nach der Kodak Grayscale: Links das hellste Exemplar (9,7), rechts das dunkelste (16), in der Mitte die häufigste Mantelfärbung (11) (© Andreas Noeske).
 |
Abb. 10: Verteilung der Kodak-Grauwerte bei Amrumer Heringsmöwen Larus fuscus. 74,2% der Vögel liegen im Bereich 11-13, der von Jonsson (1998), Malling Olsen & Larsson (2003) und Howell & Dunn (2007) als Variationsbreite von intermedius angegeben wird.
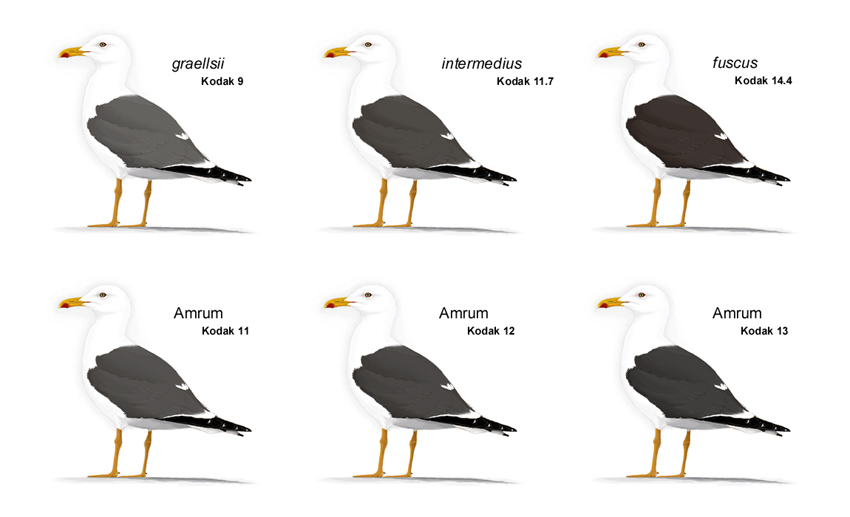
Abb. 11: Amrumer Heringsmöwen Larus fuscus mit den drei Kodak-Grauwerten, die 74,2% der Vögel zeigen, im Vergleich zu Vögeln, welche die durchschnittliche Mantelfärbung der jeweiligen Unterart aufweisen (© Andreas Noeske).
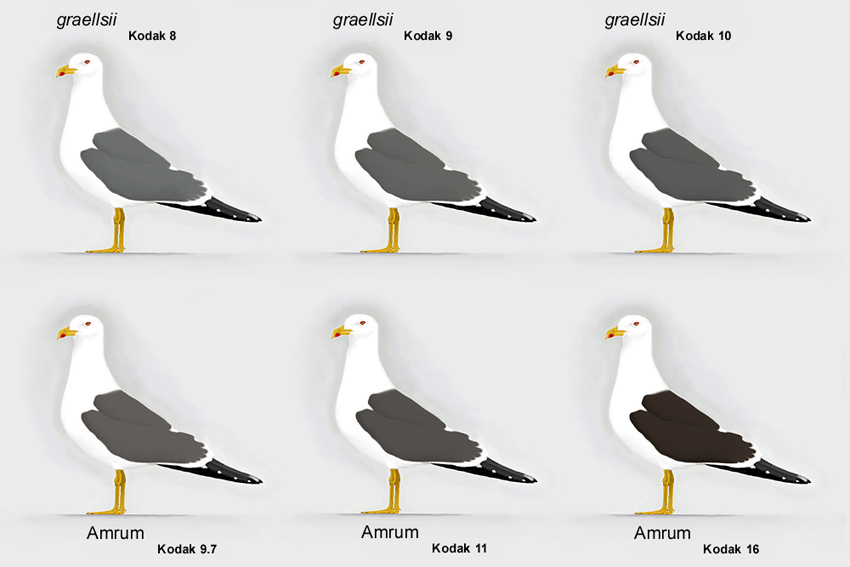
Abb. 12: Variationsbreite bei Amrumer Heringsmöwen Larus fuscus im Vergleich zu graellsii (© Andreas Noeske).
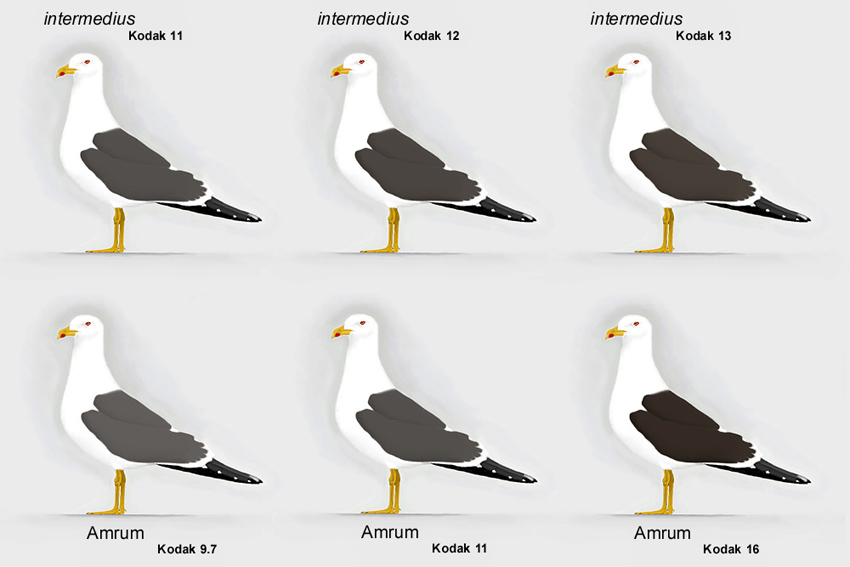
Abb. 13: Variationsbreite bei Amrumer Heringsmöwen Larus fuscus im Vergleich zu intermedius (© Andreas Noeske).
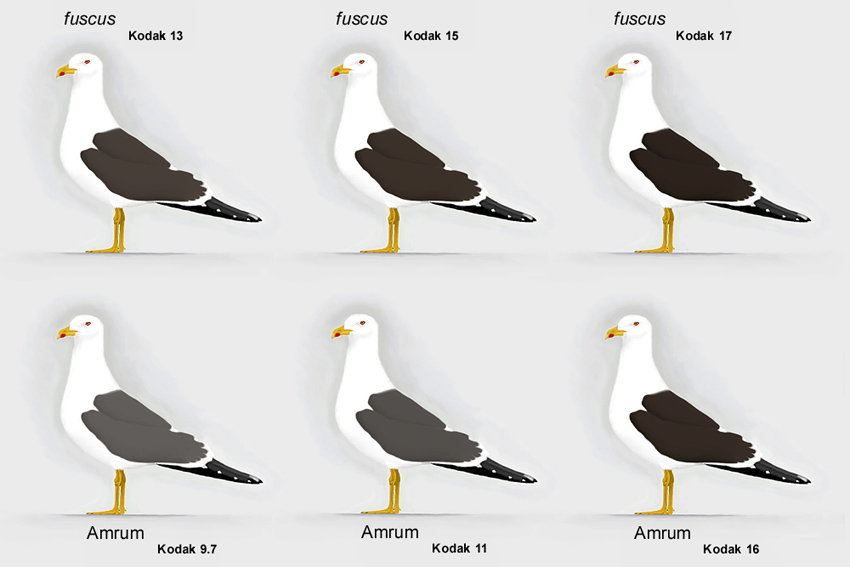
Abb. 14: Variationsbreite bei Amrumer Heringsmöwen Larus fuscus im Vergleich zu fuscus (© Andreas Noeske).
Die im Durchschnitt etwas schlankere, langflügeligere und insgesamt zierlichere Gestalt von fuscus ist in dieser Abbildung nicht berücksichtigt. Siehe hierzu Abb.15, 31, 33.
 |
Abb. 15: Heringsmöwe Larus fuscus fuscus, Tampere, Finnland, 2. Mai 2004 (© Visa Rauste).
Farbringkombination: C1AK
Beringt am 1. Juli 1999, nicht flügge, Kangasala/Tampere, Finnland (61° 28' N, 24° 04' E, Google Maps).
Amrumer Grauwerte nach Munsell
Die Munsell-Grauwerte liegen bei graellsii zwischen 3,3-4,1 (Mittelwert = 3,7), bei fuscus zwischen 1,9-2,8 (Mittelwert = 2,3) und bei intermedius zwischen 2,1-3,5 (Mittelwert = 3,0) (Barth 1966, 1968; Glutz von Blotzheim & Bauer 1982).
Die Mantelfärbung von Amrumer Heringsmöwen variiert zwischen 1,75-3,5. Der Median beträgt 3,0, der häufigste Grauwert ist ebenfalls 3,0. Die Variationsbreite wird im Vergleich der Federn mit den Grautafeln von Munsell (1929) fotografisch dargestellt (Abb.16).
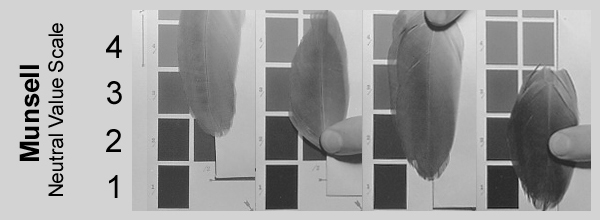 |
Abb. 16: Variationsbreite bei Amrumer Heringsmöwen Larus fuscus im Vergleich mit den Grautafeln von Munsell (1929) (© Andreas Noeske).
Die sich für Amrum ergebende Verteilung wird im Vergleich zu den anderen europäischen Populationen in Abb.17 wiedergegeben. Die geografische Verteilung des häufigsten Grauwertes einer Population wird in Abb.18 dargestellt. Tab.1 fasst die Daten der Mantelfärbung in verschiedenen Heringsmöwen-Populationen zusammen.

Abb. 17: Verteilung der Munsell-Grauwerte bei verschiedenen Populationen der Heringsmöwe Larus fuscus (erweitert nach Barth 1968). Grauwerte im Bereich 2,9-3,2 (grauer Balken) konnte Barth bei fuscus und graellsii nicht feststellen.
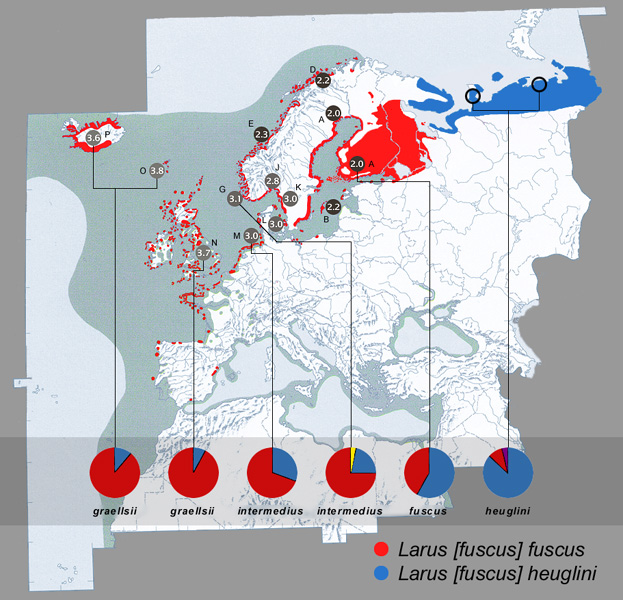
Abb. 18: Westpaläarktisches Brutgebiet der Heringsmöwen-Gruppe Larus fuscus (verändert nach Snow & Perrins 1998, Malling Olsen & Larsson 2003, lintuatlas.fi, Evgeny Potorochin pers. Mitt.). Darüber hinaus brütet die Heringsmöwe auch in W Grönland (Howell & Dunn 2007), auf den Kanaren und Spitzbergen (nach BirdLife International). Zur unterschiedlichen Auslegung des Artumfangs der Heringsmöwe siehe Yésou (2002), Sangster et al. (2007) und Collinson et al. (2008).
Dargestellt sind die geografische Variation der Mantelfärbung in Munsell-Grauwerten (Modalwert der Populationen siehe auch Tab.1, Untersuchungsgebiete verändert nach Barth 1966) und die geografische Variation der Haplotyphäufigkeiten in Kreisdiagrammen (verändert nach Liebers 2000 und Helbig et al. 2004). Weitere Ergebnisse und Interpretationen finden sich auch in Liebers & Helbig (2002): Phylogeography and colonization history of Lesser Black-backed Gulls (Larus fuscus) as revealed by mtDNA sequences).
Tab. 1: Mantelfärbungen bei verschiedenen Populationen der Heringsmöwe Larus fuscus in Munsell-Grauwerten. Angegeben sind Stichprobenumfang (n), Mittelwert, Median, Modalwert, Variationsbreite und Verteilung nach der Helligkeit des Mantels in Prozent (erweitert nach Barth 1968). Bei der Verteilung von schwarzen, dunkelgrauen und hellgrauen Heringsmöwen ist zu berücksichtigen, dass Barth den intermediären Bereich auf 2,7-3,2 festgelegt hat, was für diese Tabelle übernommen wurde.
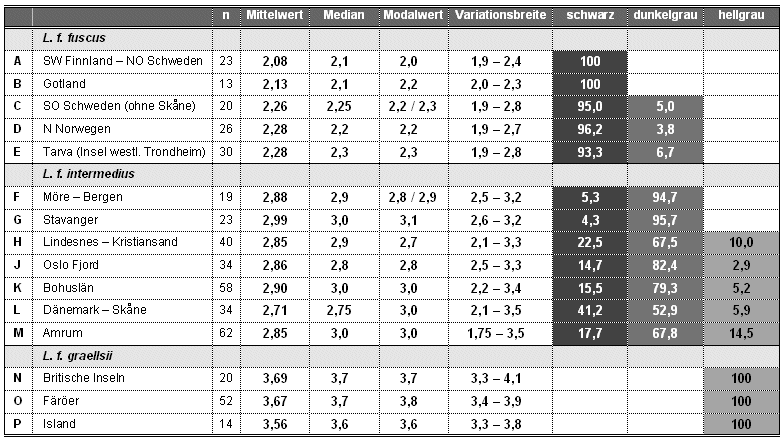 |
 |
Abb. 19: Heringsmöwe Larus fuscus, Helgoland, 20. Juni 2006, 20:36 Uhr (© Martin Gottschling).
 |
Abb. 20: Heringsmöwe Larus fuscus, Helgoland, 1. Juni 2007, 20:20 Uhr (© Jochen Dierschke).
 |
Abb. 21: Heringsmöwe Larus fuscus, Helgoland, 16. April 2006, 15:43 Uhr (© Martin Gottschling).
 |
Abb. 22: Heringsmöwe Larus fuscus, Helgoland, 20. Juni 2006, 19:50 Uhr (© Martin Gottschling).
 |
Abb. 23: Heringsmöwen-Paar Larus fuscus, Helgoland, 9. Juni 2006, 16:16 Uhr (© Martin Gottschling).
Anmerkung zur Geschlechtsbestimmung: Der Sexualdimorphismus bei Lariden äußert sich in der Körpergröße. Die Männchen sind bei einem erheblichen Überschneidungsbereich der Variationsbreiten durchschnittlich größer. Eine Geschlechtsbestimmung im Feld ist aber nur unter günstigen Umständen möglich, wenn man beide Partner eines Paares in Nestnähe nebeneinander sieht. Das Männchen ist immer größer als seine Partnerin. Die Bestimmung ist dann eindeutig, obwohl der Unterschied manchmal kaum zu erkennen ist. Bei diesem Paar ist der Größenunterschied am Schnabel besonders auffällig. Bestes Einzelmerkmal zur Trennung von Männchen und Weibchen ist die Schnabel-Kopf-Länge (Coulson et al. 1983).
Diskussion
So seltsam es im 21. Jahrhundert auch klingen mag, auch heute ist unter den Experten noch kein Konsens gefunden worden, welche Taxa der Heringsmöwe zugeordnet werden sollten. Im Gegensatz zu Helbig (2005) und Barthel & Helbig (2005), die das Superspezieskonzept anwenden, wird die Heringsmöwe von Sangster et al. (2007) und Collinson et al. (2008) weiter gefasst und umfasst zusätzlich noch die Formen heuglini, taimyrensis und barabensis. Die Tundramöwe L. fuscus heuglini wird hier also lediglich als Unterart der Heringsmöwe gesehen und nicht als stärker differenzierte Allospezies L. [fuscus] heuglini (oder kurz L. heuglini) einer Superspezies L. [fuscus] mit - im Gegensatz zu Unterarten - eingeschränkterem Genfluss. Dieses Umsortieren beruht nach den Informationen in Sangster et al. (2007) nicht auf neuen Erkenntnissen, sondern auf einer anderen Interpretation der auch Helbig (2005) vorliegenden Arbeiten. Darüber hinaus ist zu berücksichtigen, dass die Anordnung der Großmöwenformen auch auf verschiedenen Auslegungen unterschiedlicher Artkonzepte (Helbig 2000) beruht. Während Helbig für eine konsequente Anwendung des um den Superspeziesbegriff erweiterten Biologischen Artkonzepts plädierte, war/ist Sangster ein Vertreter des Phylogenetischen Artkonzepts.
Auch wenn es heute gesichert erscheint, dass wir graellsii, intermedius und fuscus als Heringsmöwe im engeren Sinn ansehen dürfen, so wurde selbst das bezweifelt. Denn in den Niederlanden wurde die Frage nach der Unterartzugehörigkeit der dort brütenden Heringsmöwen eine Zeit lang völlig anders beurteilt. Nach Vorstellungen der "Commissie Systematiek Nederlandse Avifauna" (CSNA) der Dutch Birding Association war eine Zuordnung dieser Heringsmöwen weniger kompliziert, denn nach dem dort durch einzelne Autoren in Mode gebrachten Phylogenetischen Artkonzept sollte es sich bei Larus graellsii (Lesser Black-backed Gull) und Larus fuscus (Baltic Gull) um zwei verschiedene Arten handeln. Da es nach diesem Konzept keine Unterarten gibt, war das Problem bisheriger Autoren und Beobachter scheinbar gelöst. Es bleibt allerdings zu bedenken, dass Sangster et al. (1998) - ohne eigene Ergebnisse zu liefern - lediglich bereits vorliegendes Material zu Unterschieden in Morphologie, Mauser, Nahrungserwerb und Brutverhalten mangelhaft interpretiert und Unterschiede überstrapaziert haben. Eine nachvollziehbare Analyse niederländischer Kolonien und die Einordnung der Brutvögel im Vergleich mit den Nachbarpopulationen fehlen damit nach wie vor. „Es kommt bei weiträumig und insbesondere kontinuierlich verbreiteten Arten in erster Linie auf die Analyse der regionalen Muster der geographischen Variation an und nicht auf eine bloße taxonomische Unterteilung dieser Arten in Subspezies" (Haffer 1985) oder das Splitten in Phylogenetische Arten. Dieser Ansatz der CSNA wurde im übrigen europäischen Raum daher auch nicht nachvollzogen (AERC TAC 2003) und in Holland dann wieder verworfen (Sangster et al. 2003).
Der Artstatus von graellsii (inkl. intermedius) wird durch Untersuchungen zur genetischen Differenzierung der Heringsmöwen i.w.S. nicht gestützt (Crochet et al. 2002, Liebers & Helbig 2002, Helbig et al. 2004). Die fuscus- und graellsii-Populationen zeigen denselben Haplotyp. Die geografische Variation der Haplotyphäufigkeiten bei graellsii, intermedius und fuscus verläuft dabei klinal etwa parallel zur Mantelfärbung (Abb. 18). Die mtDNA-Unterschiede zwischen fuscus aus SW Finnland und intermedius sind nur gering ohne Hinweise auf reproduktive Barrieren. Insgesamt gibt es keine Anzeichen für eingeschränkten Genfluss zwischen fuscus und graellsii. Das Fehlen von Haplotypunterschieden deutet darauf hin, dass die Trennung zwischen fuscus und graellsii ein wesentlich jüngeres Ereignis ist als die Trennung zwischen guten Arten der Großmöwengruppe.
Was ist eine Unterart?
Als Subspezies werden phänotypisch ähnliche Gruppen von Populationen einer Art zusammengefasst, die ein geographisches Teilgebiet des Artareals bewohnen und sich taxonomisch von anderen solchen Populationsgruppen der Art unterscheiden (Mayr 1975, Glutz von Blotzheim & Bauer 1985). Im Gegensatz zur Biologischen Art, die eine real existierende, ökologisch segregierte und reproduktiv isolierte Einheit der Natur und daher objektiv begrenzt und erkennbar ist, sind Unterarten Kunstprodukte des menschlichen Ordnungsbedürfnisses, die durch Mittelwertsunterschiede und Standardabweichungen unscharf und willkürlich gegeneinander abgegrenzt werden. Dabei bleibt es im Ermessen des Taxonomen, was er für unterscheidbar und taxonomisch relevant hält. Aufgrund fehlender Genflußbarrieren und „Unterartgrenzen" zwischen den Populationsgruppen sind Übergangspopulationen mit sukzessiven (klinalen) Merkmalsänderungen eher die Regel als eine Ausnahme. Auch Populationen an den Enden eines Merkmalsgradienten/einer Kline sollten selbst bei voller Diagnostizierbarkeit (z.B. Mantelfärbung bei fuscus und graellsii) nicht als Arten angesehen werden, da der Genfluß nur durch die Entfernung, nicht aber durch intrinsische Barrieren eingeschränkt ist. Solche diagnostizierbaren Populationen rechtfertigen eine Benennung als Unterart (Helbig et al. 2002). Während jede Rosenmöwe aufgrund der arttypischen autapomorphen Merkmale jederzeit und an jedem Ort diagnostizierbar und eindeutig dem Taxon Rhodostethia rosea zuzuordnen ist, gilt dies nicht für jede Heringsmöwe bei der Zuordnung zu fuscus, intermedius oder graellsii. Ohne einen zeitlichen und räumlichen Zusammenhang sind viele Heringsmöwen nicht einem der drei Taxa zuzuordnen.
Zur praktischen Abgrenzung von Unterarten
Um zu starker taxonomischer Willkür durch eine Überbetonung lokaler Unterschiede nomenklatorische Grenzen zu setzen, hat Amadon (1949) mit der sogenannten 75%-Regel ein sinnvoll erscheinendes Maß zur Abgrenzung von Unterarten vorgeschlagen, das weite Anerkennung gefunden hat (Patten & Unitt 2002, Haig et al. 2006). „Danach wird eine Population als gültige Unterart gewertet, wenn 75% der Individuen sich von „allen" (= 97%) Individuen einer bereits aufgestellten Unterart unterscheiden. Im Schnittpunkt der beiden Kurven, für die dies zutrifft, werden rund 90% der Individuen aus einer Population A von rund 90% der Individuen der Population B verschieden sein (bezogen auf eine symmetrische Verteilung)" (Mayr 1975).
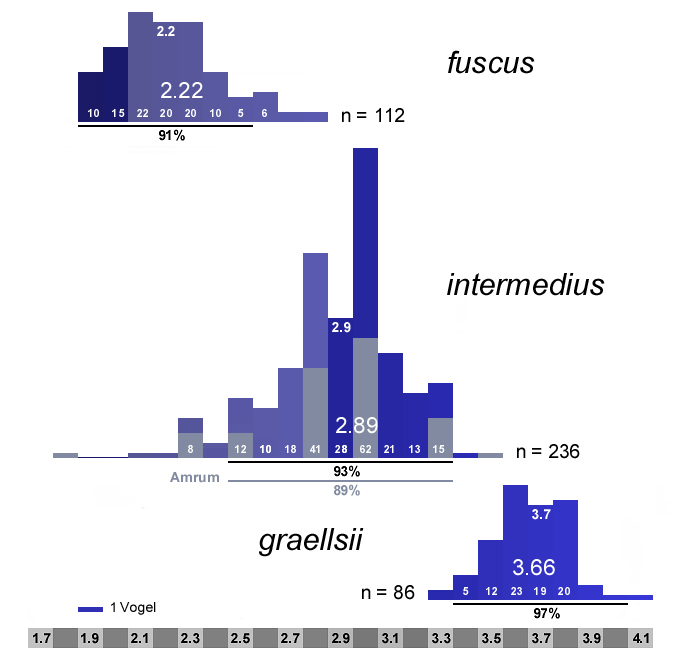 |
Abb. 24: Verteilung der Mantelfärbungen in Munsell-Grauwerten bei den drei Unterarten der Heringsmöwe Larus fuscus, Populationen zusammengefasst nach Abb.17. Bei intermedius wurden die Daten von Barth (1966) für den Bereich von der Westküste Dänemarks bis Bornholm (Gebiet L, Tab.1) nicht berücksichtigt, da der östliche Teil vom Öresund bis Bornholm nach Glutz von Blotzheim & Bauer (1982) und Jonsson (1998) zu fuscus gehört. Der Amrumer Anteil an intermedius ist hellgrau eingefärbt. Zur optischen Anpassung wurden die Amrumer Werte (1,75; 2,25; 2,75; 3,25) in dieser Darstellung aufgerundet.
Angegeben sind Stichprobenumfang (n), Anzahl der Vögel pro Grauwert, Mittelwert, Median (hochgestellt) und Grauwertbereich (schwarzer Querbalken bzw. hellgrau für Amrum), in den etwa 90% der Vögel fallen (Scheitelpunkt der Kurve beim Median). Der Überschneidungsbereich von intermedius/fuscus ist hellblau eingefärbt.
Obwohl auch statistische Verfahren letztlich nicht über die Willkür und Künstlichkeit bei der Abgrenzung von Unterarten hinwegtäuschen können, lassen sich die Unterarten der Heringsmöwe nicht nur phänotypisch sondern auch genotypsich gut begründen. Die drei Unterarten sind mitochondrien-genetisch signifikant voneinander verschieden, wobei intermedius der Nominatform fuscus sogar näher steht als graellsii. Dies spricht auch klar gegen den Versuch, fuscus unter Ausschluss der beiden anderen Taxa Artrang zuzuweisen, da die Gruppierung intermedius/graellsii paraphyletisch wäre (Liebers 2000).
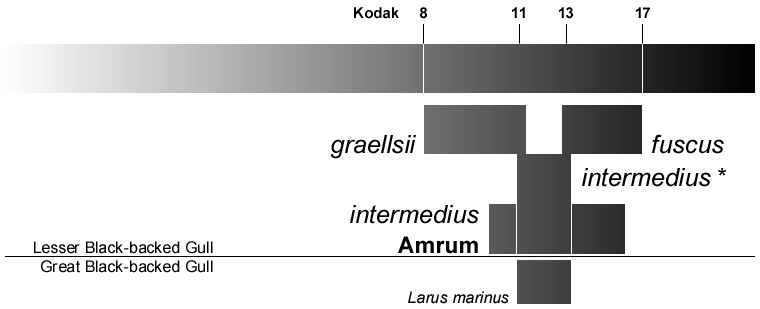 |
Abb. 25: Variationsbreite der Mantelfärbung bei der Heringsmöwe Larus fuscus mit den Graubereichen, die Jonsson (1998), Malling Olsen & Larsson (2003) und Howell & Dunn (2007) nach der Kodak-Skala für die drei Unterarten angeben.
Die Abgrenzung von intermedius* schließt nach diesen Autoren lediglich die Lücke zwischen graellsii und fuscus und gibt die wahren Verhältnisse falsch wieder. Hiernach entsteht eher der Eindruck einer starken Orientierung an starren Museumsschubladen als an lebenden Populationen. Die bereits von Barth (1966, 1968) und Glutz von Blotzheim & Bauer (1982) erwähnte große Variabilität (Munsell-Werte für L. f. intermedius = 2,1-3,5) wird von diesen Autoren ignoriert. Dagegen wird die tatsächliche Variationsbreite einer intermedius-Population durch die Amrumer Vögel angemessen wiedergegeben. Vögel, deren Mantelfärbung außerhalb der von diesen Autoren angegebenen Variationsbreite liegen, können in der Deutschen Bucht nicht als graellsii oder fuscus bestimmt werden.
In Mischgebieten haben Vögel mit z.B. einer für intermedius typischen Mantelfärbung nicht zwangsläufig auch alle anderen Merkmale, die man normalerweise mit intermedius assoziiert. Vielmehr entwickelt sich bei einer Population, die mit benachbarten Populationen im Genaustausch steht, ein Merkmalsmosaik, so dass z.B. bei den „Dutch intergrade"-Kolonien die Merkmale in der unterschiedlichsten Weise kombiniert sein können und auch in den intermedius-Kolonien Amrums und Südnorwegens Vögel auftreten, die phänotypisch von fuscus oder auch von graellsii nicht zu unterscheiden sind.
"Dutch intergrade"
Die AERC (ohne Datum) zählt die Niederlande zum Verbreitungsgebiet von intermedius, während andere Autoren die holländischen Vögel zu graellsii stellen (Liebers 2000, Liebers & Helbig 2002, Helbig et al. 2004, Muusse et al. 2005). Nach Malling Olsen & Larsson (2003) ist der subspezifische Status u.a. auch in den Niederlanden aufgrund der weitreichenden klinalen Variation unsicher. Im Vergleich zu Amrum ist eine Verschiebung zu helleren Grauwerten deutlich, die den Einfluß britischer Heringsmöwen (BTO-Ringnachweise, http://www.gull-research.org/miscellaneous/lbbgbto.html) auf holländische Kolonien anzeigt. Auch wenn nach den Haplotyp-Häufigkeiten die holländischen Heringsmöwen den britischen graellsii am nächsten stehen (Liebers & Helbig 2002), so sind sie im Durchschnitt etwas dunkler. Der Begriff „Dutch intergrade" ist eine sprachliche Hilfskonstruktion, die die Situation in Holland vermutlich am besten beschreibt, denn die Zuordnung dieser Population zu graellsii oder intermedius unterstellt eine Identität mit der britischen oder südskandinavischen Population, die in diesem Übergangsbereich tatsächlich wohl nicht vorhanden ist. Die treffendste Antwort auf die Unterartfrage in den Niederlanden dürfte schon Teixeira (1979) gegeben haben. Nach seiner Meinung sind die Niederlande gleichzeitig von Westen und Nordosten besiedelt worden, wobei es im Laufe der Zeit zu einer Vermischung der graellsii- und intermedius-Vögel gekommen ist, so dass heute eine genaue subspezifische Zuordnung der Population nicht mehr möglich ist. Britische Ringvögel und der geringe aber doch vorhandene Anteil schwarzmanteliger Exemplare (Kodak 14 und 15) machen den Genaustausch in holländischen Kolonien mit anderen Populationen deutlich.
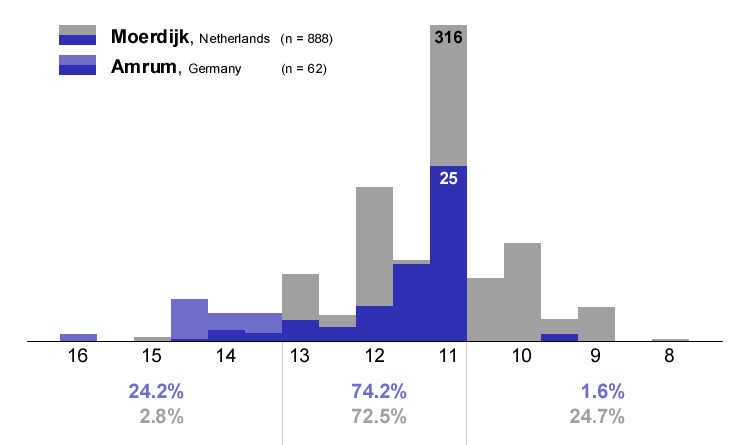 |
Abb. 26: Verteilung der Kodak-Grauwerte in einer Kolonie der Heringsmöwe Larus fuscus auf Amrum im Vergleich zur Variation in einer Kolonie bei Moerdijk/Niederlande (Daten nach Muusse et al.). Die Moerdijk-Grauwerte wurden nach der Agfa Greyscale ermittelt, die lt. Mars Muusse (in litt.) identisch mit der Kodak Grayscale sein soll. Für die vergleichende Darstellung wurden die 0,3- und 0,7-Schritte bei Amrumer Möwen zu 0,5-Werten zusammengefasst. Die Prozentzahlen geben den Anteil im jeweiligen Graubereich in den beiden Kolonien an.
 |
Abb. 27: Heringsmöwe Larus fuscus, Brutkolonie im Hafen von Zeebrügge, Belgien, 27. Mai 2005 (© Erwin van Laar).
Siehe auch http://www.birdpix.nl/album_page.php?pic_id=21172 und http://www.keeponbirding.com/foto.jsp?id=489&parentId=162.
Vergleiche mit K3 intermedius beringt in Lindesnes/S Norwegen: http://www.gull-research.org/lbbg3cyb/j4a3.htm.
Die taxonomische Stellung der Amrumer Heringsmöwen
Die Ausbreitungsgeschichte der Heringsmöwe im 20. Jahrhundert weist die mitteleuropäische Nordseeküste als Zone sekundärer Intergradation aus. Vor allem südskandinavische intermedius-, im Südwesten auch britische graellsii- und wohl auch in geringer Zahl fuscus-Vögel haben die noch zu Beginn des 20. Jahrhunderts bestehende Verbreitungslücke in einer sekundären Kontaktzone geschlossen.
Eine Charakterisierung der Amrumer Vögel nur aufgrund der Mantelfärbung würde die Verhältnisse zu sehr vereinfachen, so dass sich durch die Hinzunahme der biometrischen Daten ein vollständigeres Bild ergibt. In den Körpermaßen vermitteln die Heringsmöwen der schleswig-holsteinischen Westküste zwischen der britischen graellsii- und der südskandinavischen intermedius- Population. Deutlicher fallen die Unterschiede zu norwegischen Vögeln der Nominatform aus (Noeske 1989). Hiernach ergibt sich für die Heringsmöwen Amrums das Bild einer heterogen zusammengesetzten Population, die Merkmale aller drei Unterarten in sich vereinigt. Der von Terschelling stammende Vogel (Abb.28) und der hohe Anteil sehr dunkler Heringsmöwen machen die unterschiedlichen Einflüsse auf die deutsche Population deutlich.
Nach der Mantelfärbung sind die Heringsmöwen an der deutschen Nordseeküste in die intermedius-Population einzuordnen, was den Ursprung dieser Brutvögel am besten zu beschreiben vermag. Dieses Ergebnis ist deckungsgleich mit den Ergebnissen von Liebers (2000) auf der mitochondrien-genetischen Ebene.
„Mitochondrien-genetisch sind alle fünf [Anm.: Heringsmöwen-] Taxa nah verwandt, aber signifikant voneinander differenziert … Der Populationsstammbaum zeigt eine Zweiteilung in „westliche" und „östliche" Heringsmöwen … In der Zusammensetzung der Haplotypen gibt es keine Unterschiede zwischen den drei graellsii-Populationen. Intermedius steht der Nominatform fuscus näher als graellsii" (Liebers 2000).
 |
Abb. 28: Große Flügeldecken im Vergleich mit den Grautafeln von Munsell (1929) (© Andreas Noeske), beurteilt als Munsell 2,9 (1986), als Munsell 3,0 bzw. Kodak 11,7 (2007): Ringnummer Arnhem 5105925, beringt am 12. Juli 1976, nicht flügge, Tweede Duintjes, Terschelling, Holland (53° 25' N, 05° 28' E, Google Maps), gefangen am 2. Juni 1986, brütend, NSG Amrum-Odde, Deutschland (54° 42' N, 08° 21' E).
Von Olaf Ekelöf (pers. Mitt.) wurden auch unter den Brutvögeln auf der Helgoländer Düne drei Heringsmöwen mit Arnhem-Metallringen beobachtet, von denen folgende Rückmeldung vorliegt: Ringnummer Arnhem 5348046, beringt am 5. Juli 2001, nicht flügge, Westerduinen, Texel, Holland (53° 03' N, 04° 43' E, Google Maps), Ring abgelesen am 13. Mai 2007 als Brutvogel, Helgoland, Deutschland (54° 11' N, 07° 55' E, Google Maps).
Im Mai 2007 konnte auch ein brütender Vogel mit Madrid-Metallring ablesen werden. Überwiegend handelte es sich aber bei den beringten Vögeln um solche, die in den Jahren zuvor auch auf der Düne beringt worden waren. Bei einem weiteren Altvogel mit blauem Codering bleibt unklar, ob es sich um einen belgischen Brutvogel gehandelt hat (Olaf Ekelöf pers. Mitt.): Ringnummer L904895, rechts dunkelblauer Farbring mit weißem Code DC.AF, beringt am 12. Juni 2006, adult, Zeebrügge, Belgien (51° 21' N, 03° 11' E, Google Maps), beobachtet am 20. Mai 2007, Helgoland, Deutschland (54° 11' N, 07° 55' E).
Der Status von fuscus- und graellsii-Vögeln an der deutschen Nordseeküste
 |
Abb. 29: Heringsmöwe Larus fuscus, Helgoland, 3. Juni 2007, 11:09 Uhr (© Stefan Pfützke).
Während sich in den Niederlanden britische graellsii als Brutvögel angesiedelt haben, fehlen solche Nachweise bislang aus deutschen Möwenkolonien. Vögel, die so hellgrau erscheinen wie in dieser Abbildung, wurden und werden immer noch als graellsii bestimmt. Dabei ist das Überqueren der Nordsee durch britische Vögel eine Ausnahme. Nach Ringfunden beurteilt treten britische Heringsmöwen in Deutschland wohl etwa so selten auf wie nordskandinavische fuscus in der Deutschen Bucht (Glutz von Blotzheim & Bauer 1982, Wernham et al. 2002). Ohne Ring sind britische graellsii in Deutschland nicht nachweisbar.
Unter den Vögeln der Amrum-Odde fällt ein Exemplar mit tief schwarzem Mantel auf, das mit einem Grauwert von 1,75 bzw. 16 außerhalb der bisher beschriebenen Variationsbreite von intermedius liegt und zu den dunkelsten Heringsmöwen überhaupt gehört. Eine solche Mantelfärbung wurde bisher nur bei Heringsmöwen der Nominatform festgestellt. Diese „schwarzen" Vögel haben daher wiederholt zu der Ansicht geführt, dass sie nordskandinavischer oder baltischer Herkunft sind.
 |
Abb. 30: Brutpaar der Heringsmöwe Larus fuscus in der Kolonie der Amrum-Odde im Mai/Juni 1986 (© Andreas Noeske).
Das kleinere, schwarzmantelige Weibchen ist von fuscus-Vögeln aus dem baltischen Raum auch strukturell nicht zu unterscheiden. Für diesen Vogel ist nicht auszuschließen, dass er aus weit nördlicher oder östlicher gelegenen Gebieten stammt.
Vergleiche mit Abb.31 und mit http://www.xs4all.nl/%7edaarruud/fuscus3a.html.
 |
Abb. 31: Heringsmöwe Larus fuscus fuscus, Tampere, Finnland, 2. Mai 2004 (© Visa Rauste).
Farbringkombination: C1C2
Beringt am 30. Juni 1996, nicht flügge, Kangasala/Tampere, Finnland (61° 28' N, 24° 04' E, Google Maps).
Vergleiche mit Abb.30.
Jonsson (1998) hat zu einer möglichen Unterscheidung von fuscus und intermedius eine erste Arbeitshypothese formuliert. Danach sollen adulte fuscus im Zeitraum Ende August/September anhand der folgenden Merkmalskombination zu bestimmen sein: Schwarzer Mantel mit 50% oder mehr bräunlich-schwarzen Schulterfedern, weißer, ungestrichelter Kopf und unvermauserte innere Handschwingen. Während Jonsson dies damals noch für ausreichend hielt, wird durch neuere Arbeiten die Eignung dieser Merkmalskombination in Frage gestellt oder sogar als ungenügend betrachtet (Adriaens 2002, AERC TAC 2003, Gibbins 2004, Muusse et al. 2005, Gibbins & Baxter ohne Datum, Muusse et al. ohne Datum). Zur Unterscheidung gegenüber intermedius, die eine sehr ähnliche Merkmalskombination zeigen können, werden derzeit aber Merkmale herangezogen, die bisher überhaupt nicht quantifizierbar sind. Das folgende Denkmodell soll daher ungefähr den quantitativen Rahmen zum Auftreten von fuscus-Vögeln an der deutschen Nordseeküste abstecken. Obwohl es teilweise bereits publizierte quantitative Angaben einbezieht, enthält es viele unbekannte Faktoren, die in diesem Unschärfebereich wirksam sind.
Ringfunde finnischer fuscus zeigen, dass <1% dieser Population im Herbst eine südwestliche Zugrichtung einschlägt (Visa Rauste in Steffen 2004, Ringfunde finnischer fuscus). Die Baltische Heringsmöwe ist schon westlich einer Linie Kiel-Genf-Marseille als sehr selten anzusehen und gilt im gesamten Nordseeraum als Ausnahmeerscheinung (Glutz von Blotzheim & Bauer 1982, Kilpi & Saurola 1983, 1984, Jonsson 1998, Steffen 2004). Da norwegische und schwedische fuscus wie finnische aus ihren Brutgebieten nach Süden bis Südosten abziehen, kann der Anteil von <1% mit westlicher Flugroute für die gesamte fuscus-Population angenommen werden. Der Gesamtbestand für fuscus wird mit 55.500 Vögeln angegeben (Wetlands International 2006, zit. in http://dda-web.de Vögel in Deutschland; Schätzung des Gesamtbestandes = Anzahl Brutpaare x 3, Simon Delany pers. Mitt.), womit sich für diese Unterart in der Deutschen Bucht im Herbst ein theoretisches maximales Potenzial von 555 fuscus-Heringsmöwen ergibt. Zur Abschätzung des Altvogelanteils werden zum einen die herbstlichen Jungvogel und der hohe Anteil immaturer Möwen bei den Ringnachweisen berücksichtigt, zum anderen der Anteil von gewöhnlich >70% Altvögeln im afrikanischen Winterquartier (Lagos/Nigeria, November bis März; Wallace zit. in Glutz von Blotzheim & Bauer 1982). Danach könnte man theoretisch im Bereich der Deutschen Bucht mit nicht mehr als 250-400 adulten/subadulten Vögeln rechnen (50-70% von 555 Heringsmöwen), die die von Jonsson (1998) für fuscus beschriebene Merkmalskombination aufweisen und tatsächlich aus dem Verbreitungsgebiet der Unterart fuscus stammen. Aufgrund der höheren Beobachtungsdichte in Westeuropa im Vergleich zu den Gebieten an der normalen östlichen fuscus-Flugroute könnte dabei der tatsächliche Anteil der westlich orientierten fuscus noch sehr viel kleiner als 1% sein. Dieses theoretische Potenzial konnte z.B. in den Niederlanden bis 2007 nur durch drei adulte fuscus bestätigt werden, die als Nestlinge in Finnland beringt wurden (Ruud Altenburg pers. Mitt.) und auch in Großbritannien mit extremer Beobachterdichte gelang der erste Nachweis von fuscus erst durch einen beringten adulten Vogel im April 2007 (Stewart 2007).
Dem stehen die deutschen und skandinavischen intermedius-Vögel gegenüber, die in der Deutschen Bucht auftreten (können) und von denen ein vermutlich nicht geringer Teil mit gleicher Merkmalskombination auszuschließen ist. Der Rastbestand dieser Unterart wird von Wetlands International (2006) mit 325.000 - 440.000 Vögeln angegeben, wovon etwa 50-70% (= 162.500 – 308.000) auf adulte/subadulte Möwen entfallen. In den Überschneidungsbereich der Mantelfärbung (2,1-2,8) zwischen fuscus und intermedius fallen bei intermedius 40,0% (= 65.000 – 123.200 Vögel, Tab.2, Abb.17). Nach Beobachtungen von Klaus Malling Olsen haben 5-10% (überwiegend dunkle intermedius) im Winterquartier (Dänemark, Großbritannien, Frankreich, Marokko) ebenso weiße Köpfe wie dies bei fuscus die Regel ist, d.h. es bleiben danach immer noch mindestens 3.250 – 6.160 Vögel (= 5%), die nach der Mantelfärbung und dem weißen Kopf nicht von fuscus zu unterscheiden sind. Größe, Struktur und Mauserzustand sind zwar weitere Indizien bei der Bestimmung von fuscus, sie sind aber keine sicheren Merkmale (Rauste 1999, Muusse et al. 2005, Muusse et al. ohne Datum, http://www.xs4all.nl/%7edaarruud/fuscus3h.html) und Gibbins (2004) meint, dass spät mausernde intermedius wahrscheinlich nicht so selten sind, wie Jonsson (1998) annimmt.
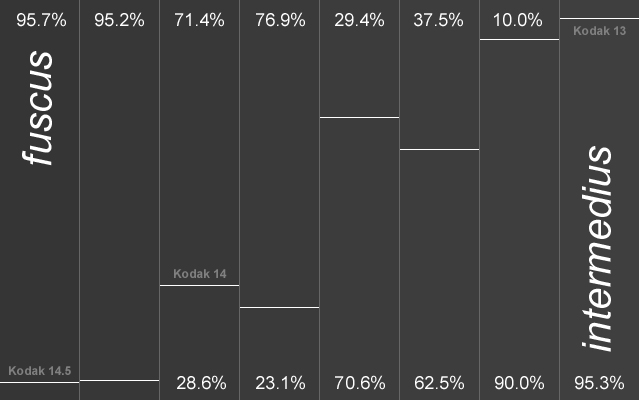 |
Abb. 32: Überschneidungsbereich der Mantelfärbung zwischen fuscus und intermedius und deren Anteil am ungefähren Graubereich. Dargestellt ist die tatsächliche Mantelfärbung im fast stufenlosen Kodak-Bereich 13-14,5. Dieser entspricht etwa dem Bereich 2,1-2,8 nach Munsell, vergleiche mit der Überschneidung zwischen Heringsmöwen von Tarva (fuscus) und Lindesnes-Kristiansand (intermedius) (Abb.17).
 |
Abb. 33: Heringsmöwe Larus fuscus fuscus, Stockholm, Schweden, 14. Juni 2006 (© Martin Lofgren).
Weitere Fotos unter http://www.wildbirdgallery.com/images/birds/larus_fuscus/fuscus.htm.
 |
Abb. 34a: Heringsmöwe Larus fuscus intermedius, Bergen, Norwegen, 2. April 2002 (© Frode Falkenberg).
Weitere Fotos unter http://cyberbirding.uib.no/gull/.
Vergleiche mit http://www.wildbirdgallery.com/images/birds/larus_fuscus/intermedius.htm.
Die relativ langen Flügel und kurzen Läufe zusammen mit der etwas geringeren Körpergröße verleihen der Baltischen Heringsmöwe eine elegante Erscheinung. Die Handschwingenprojektion von fuscus ist auffallend groß und nach Rauste (1999) "doppelt so lang wie der Überstand der längsten Schirmfeder über die Großen Armdecken". Gibbins (2004) hat erstmals versucht, diesen strukturellen Unterschied gegenüber intermedius zu quantifizieren. Nach seinen Messungen von Fotos stehender Vögel war das durchschnittliche Verhältnis Handschwingenprojektion/Länge des Tarsometatarsus (= Tarsus/Lauf, gemessen von der Mitte des Intertarsalgelenks bis zum Boden = red/yellow1 in Abb. 34b) bei fuscus 1,3 (1,0 - 1,37; n = 15), bei graellsii 0,98 (0,78 - 1,1; n = 15). Intermedius lag näher an fuscus mit einem mittleren Quotienten von 1,16 (1,1 - 1,2; n = 10). Werte > 1,2 sollten für fuscus sprechen. Aufgrund von Mauser oder Abnutzung ist die Handschwingenprojektion vieler fuscus allerdings kleiner und überlappt mit intermedius, so dass ein Vogel mit einem Wert von 1,1 - 1,2 sowohl auf fuscus als auch intermedius hindeutet. Gibbins gibt aber auch zu bedenken, dass sein Ansatz an einer größeren Stichprobe zu bestätigen wäre. Die testweise Anwendung dieses Ansatzes an einem Foto norwegischer intermedius deutet auf eine möglicherweise hohe Fehlerquote hin, denn Beobachtungen beringter Vögel zeigen, dass intermedius strukturell keineswegs einheitlich ist (Mars Muusse in Gibbins 2004).
 |
Abb. 34b: Heringsmöwe Larus fuscus intermedius, Bergen, Norwegen, 2. April 2002 (© Frode Falkenberg).
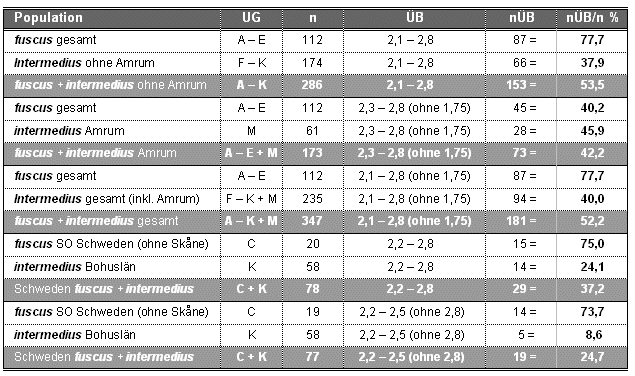
Tab. 2: Überschneidungsbereiche der Mantelfärbung zwischen verschiedenen fuscus- und intermedius-Populationen (siehe Abb.17), UG = Untersuchungsgebiete (siehe Tab.1), n = Stichprobenumfang, ÜB = Überschneidungsbereich zwischen den jeweils verglichenen Populationen, nÜB = Anzahl der Vögel der jeweiligen Population im Überschneidungsbereich, nÜB/n % = prozentualer Anteil der Vögel im ÜB im Vergleich zum Stichprobenumfang. Das Gebiet Dänemark - Skåne (L) wird bei diesem Vergleich nicht berücksichtigt, da hier keine klare Trennung zwischen dänischen intermedius und schwedischen fuscus vorliegt.
Die Angaben von Jonsson (1998) zum Überschneidungsbereich intermedius/fuscus sind nicht repräsentativ, da sie sich nur auf Teilgebiete der schwedischen West- und Ostküste beziehen. Darüber hinaus bleiben seine Berechnungen unklar, da weder Stichprobenumfänge, exakte Variationsbreiten, Überschneidungsbereich noch jeweiliger Anteil sich überschneidender Vögel bei fuscus und intermedius angegeben werden.
 |
In Westeuropa ist auch eine Kombination von schwarzem Mantel mit bräunlichem Anflug, weißem Kopf, Mauserzustand der inneren Handschwingen, nur einem weißem Subapikalspiegel auf HS 10, geringer Größe und strukturellen Merkmalen nicht diagnostisch für fuscus (Abb.35).
Dem theoretischen Potenzial von 250-400 adulten/subadulten fuscus-Vögeln könnten in der Größenordnung etwa 3.000 - 6.000 intermedius-Vögel gegenüber stehen, die nach Mantelfärbung und weißem Kopf (im Herbst/Winter) nicht von fuscus zu unterscheiden sind. Wenn Größe, Struktur und Mauserzustand als weitere Indizien diese Anzahl von fuscus- ähnlichen intermedius-Vögeln auf angenommen 10% reduzieren, läge ein theoretisch mögliches Verhältnis von adulten/subadulten fuscus- zu intermedius-Vögeln, die im Herbst an der deutschen Nordseeküste diese Merkmalskombination möglicherweise zeigen, bei etwa 1:1 (250-400 fuscus zu 300-600 intermedius). Hiernach könnte also die Irrtumswahrscheinlichkeit für die bei Steffen (2004) als fuscus-Nachweis deklarierte Beobachtung einer K3-Heringsmöwe vom 29. Oktober 2003 durchaus bei 50% liegen.
Der spekulative Rahmen in diesem Grenzbereich der Vogelbestimmung wird durch dieses Denkmodell quantitativ nur näherungsweise beschrieben. Eingedenk der Tatsache, dass der Genfluß an den von Taxonomen willkürlich definierten „Unterartgrenzen" keinen Halt macht und der Übergang von fuscus zu intermedius zumindest in der Mantelfärbung klinal ist, bleibt die Schwierigkeit bestehen, einen seriösen fuscus-Nachweis ohne Ringablesung zu erbringen. Die Niederländische Seltenheitenkommission CDNA hat auf diese Schwierigkeiten reagiert und alle angeblichen fuscus-Beobachtungen ohne Herkunftsnachweis durch einen Ring gestrichen (van der Vliet et al., Cottaar 2005) und auch das British Birds Rarities Committee BBRC geht nach dem gegenwärtigen Kenntnisstand davon aus, dass fuscus im Feld als vagrant in Westeuropa nicht bestimmbar ist, obwohl die Jonsson-Kriterien im Baltikum durchaus zur Anwendung kommen können (Colin Bradshaw pers. Mitt.). In Übereinstimmung mit der oben kalkulierten hohen Fehlerquote meinte schon Gibbins (2004), dass die Bestimmung von K3 fuscus eher Ansichtssache ist.
 |
Abb. 35: K3 Heringsmöwe Larus fuscus intermedius, Westkapelle, Holland, 2. September 2003 (© Mars Muusse).
Ringnummer: NOS 4217224
Beringt am 19. Juli 2001, nicht flügge, Hellersoy, Sogne, Vest-Agder, S Norwegen (58° 01' N, 07° 49' E, Google Maps).
Auf Populationsebene lassen sich zwar unter anderem Unterschiede in der Mantelfärbung, Größe, Struktur und Mauser ausmachen, für eine Zuordnung von Einzelvögeln ist auch weiterhin eine Kombination mehrerer Merkmale nicht ausreichend, um adulte fuscus und intermedius oder intermedius und graellsii zu unterscheiden. Die Bestimmung unberingter britischer graellsii oder baltischer fuscus an der deutschen Nordseeküste bleibt weiterhin hochgradig spekulativ.
Weitere Fotos dieses Vogels unter http://www.gull-research.org/lbbg3cyc/nos4217224.htm, vergleiche mit http://www.xs4all.nl/%7edaarruud/fuscus3b.html.
An der deutschen Nordseeküste ist die Zuordnung einzelner Vögel mit besonders dunklem oder sehr hellem Mantel zur baltischen bzw. britischen Population auch in Kombination mit weiteren Merkmalen aufgrund der Variation der Merkmalskombinationen innerhalb der Taxa weiterhin unmöglich. Viele Beobachter werden durch die oft als quälend empfundene diffizile Möwenbestimmung ohnehin bis über ihre Grenzen gefordert und die enorme Variabiliät der verschiedenen Kleider mit unzähligen Ausnahmen sprengt dabei wohl bei den meisten die Aufnahmefähigkeit und Nachvollziehbarkeit. Da kommt es als willkommener Trost, wenn selbst Experten meinen, dass das wohl beste Bestimmungskriterium für die in Westeuropa seltenen fuscus-Vögel der richtige Ring ist (AERC TAC 2003, Muusse et al. 2005, Gibbins & Baxter).
Danksagung
Ich bedanke mich besonders beim Verein Jordsand, bei Prof. Dr. D. Mossakowski und Dr. G. Vauk, ohne deren Unterstützung diese Arbeit nicht möglich gewesen wäre. Stefan Pfützke war mir auf verschiedene Weise besonders behilflich. Ralf Aumüller, Jochen Dierschke, Martin Gottschling, Axel Halley und Thorsten Krüger machten letzte Verbesserungsvorschläge am Manuskript. Christoph Zöckler war freundlicherweise beim englischen Text behilflich. Darüber hinaus bedanke ich mich für ihre Unterstützung bei Sven Achtermann, Peter Adriaens, Ruud Altenburg, Colin Bradshaw, Andreas Buchheim, Simon Delany, Volker Dierschke, Ralf Eikhorst, Olaf Ekelöf, Frode Falkenberg, Miel Ferdinande, Bernd-Olaf Flore, Chris Gibbins, Jukka Haapala, Jürgen Haffer, Susan Haig, Risto Juvaste, Alex Kaat, Antero Lindholm, Martin Lofgren, Pat Lonergan, Carl Erik Mabeck, Klaus Malling Olsen, Mars Muusse, Hans Wolfgang Nehls, Evgeny Potorochin, Laurent Raty, Visa Rauste, Gabriel Schuler, Joachim Seitz, Dietrich Sellin, Rainer Sottorf, Konrad Thye, Norbert Uhlhaas und Erwin van Laar.
Literatur
- Adriaens, P. 2002: Some confusing Lesser Black-backed Gulls (Larus fuscus). http://www.surfbirds.com/mb/Features/gulls/LBB/lbb-gull.html
- AERC TAC 2003: AERC TAC's Taxonomic Recommendations (1 December 2003). Pending, S.12-13. http://www.aerc.eu/aerc_tac.htm
- AERC ohne Datum: AERC TAC Checklist of bird taxa occurring in Western Palearctic region, with distributional notes on subspecies - 15th Draft. S.20. http://www.aerc.eu/DOCS/Bird_taxa_of%20_the_WP15.pdf http://www.aerc.eu/Documents.htm
- Barth, E.K. 1966: Mantle colour as a taxonomic feature in Larus argentatus and Larus fuscus. Nytt Mag. Zool. 13: 56-82.
- Barth, E.K. 1967: Standard body measurements in Larus argentatus, L. fuscus, L. canus, and L. marinus. Nytt Mag. Zool. 14: 7-83.
- Barth, E.K. 1968: The circumpolar systematics of Larus argentatus and Larus fuscus with special reference to the Norwegian populations. Nytt Mag. Zool. 15, Suppl. 1: 1-50.
- Barth, E.K. 1975: Taxonomy of Larus argentatus and Larus fuscus in north-western Europe. Ornis Scand. 6: 49-63.
- Barthel, P.H. & A.J. Helbig 2005: Artenliste der Vögel Deutschlands. Limicola 19: 89-111.
- Bauer, H.-G., E. Bezzel & W. Fiedler 2005: Das Kompendium der Vögel Mitteleuropas. Bd. 1. 2. Aufl. Wiebelsheim.
- Busch, F.-D. 1974: Heringsmöwe (Larus fuscus) 1973 auf der Insel Spiekeroog brütend. Vogelk. Ber. Niedersachs. 6: 18.
- Busche, G. & R.K. Berndt 1982: Ornithologischer Jahresbericht für Schleswig-Holstein 1980. Corax 9: 9-37.
- Collinson, J.M., D.T. Parkin, A.G. Knox, G. Sangster & L. Svensson 2008: Species boundaries in the Herring and Lesser Black-backed Gull complex. Brit. Birds 101: 340-363.
- Cottaar, F. 2005: Baltic Gull at IJmuiden in September 2002. Dutch Birding 27: 40. http://www.eurobirding.com/birdingmagazines/artinfo.php?id=7944
- Coulson, J.C., C.S. Thomas, J.E.L. Butterfield, N. Duncan, P. Monaghan & C. Shedden 1983: The use of head and bill length to sex live gulls Laridae. Ibis 125: 549-557.
- Cramp, S. & K.E.L. Simmons (Hrsg.) 1983: The Birds of the Western Palearctic. Vol. 3. Oxford, London, New York.
- Crochet, P.-A., J.-D. Lebreton & F. Bonhomme 2002: Systematics of large white-headed gulls: Patterns of mitochondrial DNA variation in western European taxa. The Auk 119: 603-620. http://www.gull-research.org/papers/21crochet.pdf
- Gibbins, C. 2004: Is it possible to identify Baltic and Heuglin's Gulls? Birding Scotland 7: 153-186. http://www.gull-research.org/papers/01gibbins.pdf (4.19 MB)
- Gibbins, C. & P. Baxter ohne Datum: Baltic and Heuglin's Gull - Photo Essay. http://www.surfbirds.com/mb/Features/gulls/baltic-heuglins-gulls.html
- Glutz von Blotzheim, U.N. & K.M. Bauer 1982: Handbuch der Vögel Mitteleuropas. Bd. 8/I. Wiesbaden.
- Glutz von Blotzheim, U.N. & K.M. Bauer 1985: Handbuch der Vögel Mitteleuropas. Bd. 10/I. Wiesbaden.
- Goethe, F. 1951: Westliche Heringsmöwe (Larus fuscus graellsii) augenscheinlich Brutvogel auf der Helgoländer Düne. Vogelwarte 16: 82.
- Goethe, F. 1957: Die Westliche Heringsmöwe als Brutvogel auf Memmert und anderen deutschen Inseln. Beitr. Naturkde Niedersachsen 10: 49-60.
- Haffer, J. 1985: Passeres oder Oscines – Singvögel. in: Glutz von Blotzheim, U.N. & K.M. Bauer: Handbuch der Vögel Mitteleuropas. Bd. 10/I. Wiesbaden. S.46-62.
- Haig, S.M. E.A. Beever, S.M. Chambers, H.M. Draheim, B.D. Dugger, S. Dunham, E. Elliott-Smith, J.B. Fontaine, D.C. Kesler, B.J. Knaus, I.F. Lopes, P. Loschl, T.D. Mullins, L.M. Sheffield 2006: Taxonomic Considerations in Listing Subspecies Under the U.S. Endangered Species Act. Conservation Biology 20: 1584–1594.
- Hanoldt, W. 1974: Heringsmöwen-Brut auf Trischen, Probleme der Rassenverbreitung. Vogel und Heimat 23: 270-271.
- Hausmann, K. 1969: Beobachtungen an der Westlichen Heringsmöwe (Larus fuscus graellsii) auf der Insel Memmert. Orn. Mitt. 21: 119-126.
- Helbig, A.J. 2000: Was ist eine Vogel-„Art"? – Ein Beitrag zur aktuellen Diskussion um Artkonzepte in der Ornithologie. Teil I. Limicola 14: 57-79.
- Helbig, A.J. 2005: Anmerkungen zur Systematik und Taxonomie der Artenliste der Vögel Deutschlands. Limicola 19: 112-128.
- Helbig, A.J., A.G. Knox, D.T. Parkin, G. Sangster & M. Collinson 2002: Guidelines for assigning species rank. Ibis 144: 518-525. http://www.blackwell-synergy.com/doi/full/10.1046/j.1474-919X.2002.00091.x
- Helbig, A.J., D. Liebers & P. de Knijff 2004: Artbildung und Verwandtschaftsverhältnisse im Silber-Heringsmöwen-Komplex Larus argentatus/fuscus. Limicola 18: 233-258.
- Howell, S.N.G. & J.L. Dunn 2007: Gulls of the Americas. Boston, New York.
- Jonsson, L. 1998: Baltic Lesser Black-backed Gull Larus fuscus fuscus - moult, ageing and identification. Birding World 11: 295-317.
- Kilpi, M. & P. Saurola 1983: Geographic distribution of breeding season recoveries of adult and immature Larus marinus, L. argentatus and L. fuscus ringed in Finland. Ornis Fennica 60: 117-125.
- Kilpi, M. & P. Saurola 1984: Migration and wintering strategies of juvenile and adult Larus marinus, L. argentatus and L. fuscus from Finland. Ornis Fennica 61: 1-8.
- Liebers, D. 2000: Phylogeographische Differenzierung und Verwandtschaftsbeziehungen von Großmöwen der Larus argentatus – fuscus – cachinnans Gruppe: Untersuchungen anhand von DNA-Sequenzen der mitochondrialen Kontrollregion. Dissertation, Universität Greifswald. http://www.greengull.de/img/dissertation_dliebers.pdf
- Liebers, D. & A.J. Helbig 2002: Phylogeography and colonization history of Lesser Black-backed Gulls (Larus fuscus) as revealed by mtDNA sequences. J. Evol. Biol. 15: 1021-1033. http://www.blackwell-synergy.com/doi/abs/10.1046/j.1420-9101.2002.00454.x
- Malling Olsen, K. & H. Larsson 2003: Gulls of North America, Europe, and Asia. Princeton, Oxford.
- Meier, O.G. & P. Todt 1985: Die Besiedlung Trischens durch Möwen (Laridae), insbesondere durch die Heringsmöwe (Larus fuscus). Dithmarschen 3: 85-86.
- Munsell, A.H. 1929: Book of color. Standard edition; Munsell Color Comp., Baltimore. Serial No. 218, Exemplar der Staatsbibliothek Berlin.
- Muusse, M., T. Muusse & B.-J. Luijendijk ohne Datum: Immature LBBGs resembling fuscus in Western Europe (so-called discusfus LBBG's). http://www.gull-research.org/indexelements/discussion_page.htm
- Muusse, T.O.V., M.J.M. Muusse, B.-J. Luijendijk & R.G.M. Altenburg 2005: Identification update: moult variability in 3rd calendar-year Lesser Black-backed Gulls. Birding World 18: 338-348.
- Noeske, A. 1989: Die systematische Stellung der an der deutschen Nordseeküste brütenden Heringsmöwen (Larus fuscus). Unveröffent. Diplomarbeit, Universität Bremen.
- Patten, M.A. & P. Unitt 2002: Diagnosability versus mean differences of Sage Sparrow subspecies. The Auk 119: 26-35. http://findarticles.com/p/articles/mi_qa3793/is_200201/ai_n9075352
- Pfeifer, G. 2003: Die Vögel der Insel Sylt. Husum.
- Rauste, V. 1999: Kennzeichen und Mauser von „Baltischen Heringsmöwen" Larus [fuscus] fuscus und „Tundramöwen" L. [fuscus] heuglini. Teil I + II. Limicola 13: 105-128, 153-188.
- Ridgway, R. 1912: Color Standards and Color Nomenclature. Washington. Exemplar des Instituts für Vogelforschung Wilhelmshaven.
- Sangster, G., C.J. Hazevoet, A.B. van den Berg & C.S. Roselaar 1998: Dutch avifaunal list: species concepts, taxonomic instability, and taxonomic changes in 1998. Dutch Birding 20: 22-32.
- Sangster, G., A.B. van den Berg, A.J. van Loon & C.S. Roselaar 2003: Dutch avifaunal list: taxonomic changes in 1999-2003. Ardea 91: 279-286. PDF-Version erhältlich von csna@dutchbirding.nl
- Sangster, G., J.M. Collinson, A.G. Knox, D.T. Parkin & L. Svensson 2007: Taxonomic recommendations for British birds: Fourth report. Ibis 149: 853-857. http://www3.interscience.wiley.com/cgi-bin/fulltext/118531152/HTMLSTART
- Steffen, B. 2004: Zwei Nachweise der Baltischen Heringsmöwe Larus fuscus fuscus auf Helgoland. Ornithol. Jber. Helgoland 14: 78-82.
- Stewart, P. 2007: The Baltic Gull in Gloucestershire - the first confirmed British record. Birding World 20: 152-153.
- Svensson, L., P.J. Grant, K. Mullarney & D. Zetterström 2000: Vögel Europas, Nordafrikas und Vorderasiens. Stuttgart.
- Snow, D.W. & C.M. Perrins (Hrsg.) 1998: The Birds of the Western Palearctic - Concise Edition. Vol. 1. Oxford, New York.
- Teixeira, R.M. 1979: Atlas van de Nederlandse Broedvogels. Deventer.
- Tinbergen, N. 1929: A breeding pair of Herring-Gull (Larus a.argentatus Pont.) x Lesser Black-backed Gull (Larus fuscus subspec.). Ardea 18: 1.
- van der Vliet, R.E., J. van der Laan & CDNA: Rare birds in the Netherlands in 2003. http://www.dutchbirding.nl/comm/cdna/2003.html
- van Dobben, W.H. 1931: Einzelheiten über Farbe und Brutbiologie der auf Terschelling nistenden Heringsmöwen (Larus fuscus subspec.). Ardea 20: 143-147.
- Voous, K.H. 1963: Geographic variation of Larus fuscus in northwestern Europe. Ardea 51: 16-24.
- Wernham, C.V., M.P. Toms, J.H. Marchant, J.A. Clark, G.M. Siriwardena & S.R. Baillie (Hrsg.) 2002: The Migration Atlas: movements of the birds of Britain and Ireland. London.
- Wetlands International 2006: Waterbird population estimates. 4th edition. Wetlands International, Wageningen, The Netherlands. http://dda-web.de/index.php?cat=Daten%20und%20Service&id=6&subid=1&ssc=1&lang=de&species=5910#
- Yésou, P. 2002: Trends in systematics: Systematics of Larus argentatus-cachinnans-fuscus complex revisited. Dutch Birding 24: 271-298.
- Zang, H., G. Großkopf & H. Heckenroth 1991: Die Vögel Niedersachsens, Raubmöwen bis Alken. Naturschutz Landschaftspfl. Niedersachs. B, Heft 2.6. Hannover.
Andreas Noeske, Bremen,